|
Jaroslav Ponec Aphyosemion i Roloffia Przepiękne afrykańskie proporczykowce z rodzaju Aphyosemion i Roloffia były zawsze ośrodkiem zainteresowań tych akwarystów, którzy postanowili hodować ryby z rodziny Cyprinodontidae. Ale nie tylko im, także ichtiologom, nie dały one spać. A to dlatego, że z punktu widzenia taksonomii stanowią jedną z najtrudniejszych grup. W rzadko której grupie nawarstwiło się tyle błędów i niejasności co w tej właśnie. Prawdopodobnie dlatego w ostatnim czasie wielu badaczy zaczęło intensywnie studiować (albo badać) afrykańskie proporczykowce. Na pierwszym miejscu wymienić trzeba duńskiego specjalistę J. J. Scheela, dalej S. Clausena, E. Roloffa, A, I. Raddę, W. Foercha i wielu innych. Ich zasługą jest wniesienie światła w systematykę obydwu rodzajów. A. Warunki ekologiczne a rytm życiowy. Warunki ekologiczne wpływają decydująco na życie organizmów. Ich znajomość jest dlatego nadzwyczaj ważna dla akwarystyki. To one wytworzyły obecny rytm życiowy, tj. tryb życia i rozmnażania, który ustalił się w wyniku długiej ewolucji. Z tego punktu widzenia dzielono dawniej proporczykowce na dwie grupy: gatunki wycierające się na dnie i gatunki wycierające się w roślinach. Klasyczny ten podział nie jest jednak całkiem ścisły, ponieważ kryterium sposobu tarła nie może być podstawą podziału tych dwóch wielkich grup ekologicznych. Istotnym kryterium podziału jest natomiast rozwój ikry, a według niego rozróżniamy: gatunki z nieprzerwanym (ciągłym) rozwojem ikry, gatunki z przerwanym rozwojem ikry (nieciągłym) i tak zwaną grupę przejściową (pośrednią), u której rozwój ikry może, ale nie musi być przerwany. 1. Gatunki z ciągłym rozwojem ikry. Gatunki te żyją w środowiskach, w których nawet w porze suchej utrzymuje się woda. Są to głównie potoki, rzeczki i zbiorniki wodne o dużym przepływie i najczęściej miękką oraz kwaśną wodą. Większość tych wód charakteryzuje się twardością do 1°n i odczynem lekko kwaśnym (pH 5,5 do 6,5). Rozwój ikry przebiega w sposób ciągły (nieprzerwany). Ryby wycierają się przeważanie na roślinach o delikatnych liściach, narybek lęgnie się z reguły po okresie 2 do 4 tygodni. 2. Gatunki z przerwanym rozwojem ikry. Przerwany (nieciągły) rozwój ikry jest konieczną reakcją na takie warunki życia, jak okresowe zanikanie wód (tzw. zbiorniki okresowe). Warunki te przetrzymuje ikra złożona w podłożu, a z niej, po pierwszych opadach lęgną się młode rybki. 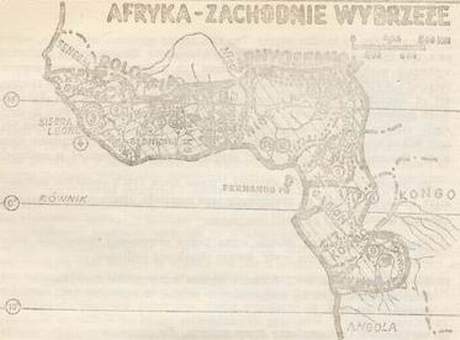 Rys. 1 Geograficzne rozmieszczenie rodzaju Aphyosemion i Roloffia. Numery grup przedstawiono w „Przeglądzie grup rodzaju Aphyosemion i Roloffia„. 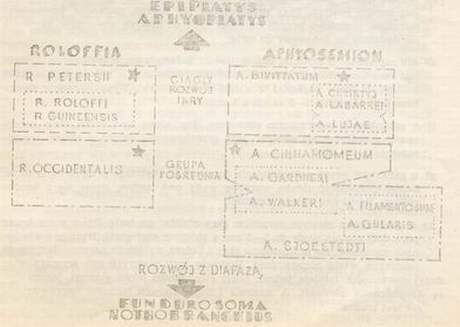 Rys. 2 Schemat pokrewieństwa filogenetycznego rodzaju Aphyosemion i Roloffia. Przedstawione są tylko typowe gatunki poszczególnych grup. Gatunki oznaczone gwiazdką mają wyższą izolację i są bardziej samodzielne niż pozostałe. Wysychanie zbiorników wodnych jest wynikiem ujemnego bilansu między parowaniem a przypływem wody w porze suchej. Jest więc logiczne, że obok siedlisk stale zalanych i zbiorników w pewnym okresie całkowicie wysychających, istnieje pośrednia grupa biotopów, których granica jest wyznaczona dwiema poprzednimi podstawowymi grupami, w których woda wysycha lub prawie wysycha na krótkie okresy. Ten stan rzeczy wywiera, oczywiście, wpływ na rytmy życiowe ryb zamieszkujących te środowiska. Gatunki grupy pośredniej wykazują większą inklinację ku jednemu lub drugiemu sposobowi rozwoju ikry, co ujawnia się różnie wewnątrz danego gatunku lub populacji, w zależności od warunków życia. Najlepszym przykładem dla tej grupy jest Aphyosemion gardneri. Czas rozwoju ikry może być bardzo zmienny, od dwu do dziesięciu tygodni. Zbyt radykalne wysuszanie zapłodnionej ikry często bywa błędem hodowlanym. Lepiej jest więc zostawić nad torfem parę milimetrów słupa wody. Czasami proporczykowce tej grupy wycierają się chętniej w roślinach. a) miękka, słabo kwaśna, odstała woda. Dobra równowaga biologiczna, zbiornik z niskim poziomem wody, dobrze obsadzony roślinami, B. Podział systematyczny rodzaju Aphyosemion i Roloffia. Jak już wspominaliśmy, systematyka dotychczasowego rodzaju Aphyosemion, który obecnie dzieli się na dwa samodzielne rodzaje Aphyosemion i Roloffia była do niedawna w niewesołej sytuacji. Ten stan był między innymi wynikiem tego, że dotychczasowa koncepcja gatunku, który był klasyfikowany przede wszystkim na podstawie wyglądu zewnętrznego, nie wystarcza. Genetyka uczy, że wygląd zewnętrzny (fenotyp) jest wynikiem możliwości określonych przez genotyp, a wykształconych pod wpływem działania warunków życia i że genotyp podlega zmienności wewnątrz samego gatunku. Dlatego punktem wyjścia jest zbadanie genotypu i założenie, że gatunki odgraniczone są od siebie izolacją reproduktywną. Znany amerykański zoolog Ernest Mayr w roku 1940 sformował koncepcję gatunku następująco: Gatunek (species) jest to grupa spokrewnionych populacji, zdolnych do wzajemnego krzyżowania się i izolowanych reproduktywnie od innych takich grup. Jeżeli tak rozumiemy gatunek, może się więc zdarzyć (a właśnie tak jest u proporczykowców), że na jednym terytorium znajduje się grupa blisko spokrewnionych populacji, które się jednak nie krzyżują między sobą i nie mają nawet potencjalnych możliwości produkowania płodnych hybrydów. Takie populacje są więc reproduktywnie izolowane, co nadaje im rangę samodzielnych gatunków. Formalnie nie mogą być one jednak nomenklaturycznie traktowane jako gatunki, ponieważ są na tyle podobne, zmienne i geograficznie wzajemnie pomieszane, że wytypowanie rozróżniających je kryteriów morfologicznych i geograficznych jest zbyt skomplikowane lub wręcz niemożliwe. Dlatego takie populacje oznaczamy tą samą nazwą gatunkową i mówimy o nadgatunku (superspecies). Nadgatunek jest więc grupą blisko spokrewnionych, ale reproduktywnie izolowanych populacji. Definicji gatunku Mayra należało koniecznie użyć w celu osiągnięcia obiektywności klasyfikacji taksonomicznej tak trudnych zbiorów jak np, rodzaj Aphyosemion. Tę nową drogą w studiach systematyki rodziny Cyprinodontidae rozpoczął duński badacz Jörgen J. Scheel, który ustalił liczbę i morfologię chromosomów, izolację reproduktywną i geograficzne rozmieszczenie poszczególnych form. Doszedł do znakomitych wyników i na jego to pracach opiera się przede wszystkim współczesna systematyka rodziny Cyprinodontidae. Tyle na razie dla zrozumienia podstawowych zasad i pojęć taksonomicznych z którymi spotkamy się w związku z rewizją obu rodzajów. Teraz już konkretnie o nowym podziale systematycznym. Dla lepszego zrozumienia wewnętrznej systematyki obu złożonych rodzajów i dla orientacji w oznaczeniu i hodowli różnych gatunków uważam za celowe rozdzielenie tych rodzajów na mniejsze zespoły, złożone z pokrewnych gatunków. Pokusiłem się na dokonanie takiego podziału, przy czym dla zespołów tych konsekwentnie, używam terminu „grupa”. Tak pojmowana „grupa” jest nieformalną jednostką taksonomiczną między gatunkiem (species) a podrodzajem (subgenus) i wyższą od nadgatunk (superspecies) W ramach klasyfikacji grup użyto równocześnie kryteriów ekologicznych, morfologicznych i geograficznych. Oznacza to, że w jednej grupie umieszczone są gatunki żyjące w podobnych warunkach ekologicznych, na zwartym obszarze geograficznym. Dla gatunków jednej grupy obowiązują wspólne zasady hodowlane. Badając pokrewieństwa poszczególnych gatunków na poziomie grup stwierdzamy, że istnieją gatunki, które ze względu na swe cechy morfologiczne nie mogą być przyłączone do żadnej z tych grup. Wydzielamy je więc w samodzielne grupy monotypowe, obejmujące jeden tylko gatunek. Jest jednak znamienne, że większość tych właśnie gatunków ma charakter nadgatunku (superspecies). Grupy noszą nazwy ich najbardziej charakterystycznego a gdzie to było możliwe znanego gatununku. Jest więc i tutaj zachowana zasada typu przyjęta w całej taksonomii. Przegląd grup rodzaju Aphyosemion i Roloffia.
Geograficzne rozmieszczenie grup przedstawione jest na rys. 1. Badając filogenetyczne pokrewieństwo afrykańskich karpieńcowatych z podrodziny Rivulinae stwierdzamy, że rodzaje Aphyosemion i Roloffia stanowią ogniwo łączące między rodzajami Epiplatys i Aphyoplatys, tj. typowymi rybami górnych warstw wody z ciągłym rozwojem ikry, a rodzajami Fundulosoma i Nothobranchius, które należą do karpieńcowatych typowych dla zbiorników sezonowych i charakteryzują się przerwanym rozwojem ikry. 1.1. R. roloffi (Ahl, 1936) – Sierra Leone (rys. 3A)  Rys. 3: Gatunki rodzaju Roloffia: A – R. roloffi, B – R. liberiensis, C – R. bertholdi, D – R. geryi, E – R. brueningi, F – R. chaytori, G – R. petersi, H – R. guineensis, J – R. occidentalis todti, K – R. occidentalis occidentalis, L – R. monroviae. Grupa ta obejmuje bardzo podobne gatunki, charakteryzujące się smukłą budową ciała, wyraźnym metalicznym ubarwieniem oraz charakterystycznym kształtem i rysunkiem płetw. R.roloffi – barwa podstawowa przeważnie zielona, płetwa ogonowa z wyraźnym żółtym obrzeżeniem. Samice mają nasadową plamę u nasady ogona.  Rys. 4: Gatunki rodzaju Aphyosemion: A – A. blvittatum, B – A. australe, C – A. ahli, D – A. calliurum, E – A. batesii, F – A. bualanum, G – A. exiguum, H – A. lujae, J – A. obseurum, K – A. santaisabelae, L – A. cameronense, M – A. cinnamomeum Ogólnie można powiedzieć, że gatunki tej grupy łatwiej jest oznaczyć na podstawie samic, niż samców. Należą do grupy ekologicznej z ciągłym rozwojem ikry, czasami jednak wycierają się na dnie i nawiązują do grupy przejściowej.  Rys. 5: Gatunki rodzaju Aphysemion: A – A. labarrei, B – A. louessense, C – A. melanopteron, D – A. christyi, E – A. cognatum, F,G,H – różne populacje A. gardneri, J – A. scheeli, K – A. ndianum, L – A. mirabilis, M – A. walkeri. 2. Grupa Roloffia petersii: 2.1. R.petersii (Sauwage, 1882)- Wybrzeże Kości Słoniowej, Ghana (rys.3G) Jest on ze wszystkich gatunków omawianego rodzaju najbardziej zbliżony do rodzaju Epiplatys. Ciało tych rybek jest brązowe w złote i zielone błyszczące plamy, z ciemnymi poprzecznymi paskami w tylnej części ciała, które są szczególnie wyraźne u młodych ryb. Nie można go pomylić z żadnym innym gatunkiem obu rodzajów. Wyciera się na roślinach, ikra rozwija się ciągle. 3. Grupa Roloffia guineensis: 3.1. R. guineensis.- (Daget, 1954) – Gwinea, Sierra Leone, Liberia (rys.3H) Różni się on od gatunków grupy R.roloffi głównie bardziej krępą budową ciała, które jest metalicznie niebiesko ubarwione i przeważnie pozbawione wyraźnych czerwonych plam. Zaokrąglona płetwa ogonowa ma w dolnej części szeroki biały lub bladożółty brzeg. Rozwój ikry przebiega ciągle.  Rys. 6: Gatunki rodzaju Aphyosemion: A – A. filamentosum, B – lokalna forma A. filamentosum, C – A. arnoldi, D – A. gulare, E – A. sjoestedti. 4. Grupa Roloffia occidentalis occidentalis (Clausen, 1965) Sierra Leone (rys. 3K) 4.1.2. Roloffia occidentalis todti (Clausen, 1965) – Sierra Leone (rys. 3J) R.occidentalis znany jest u nas jako proporczykowiec złoto-obrzeżony. Jest on krępą rybą o charakterystycznym ubarwieniu ciała. Podgatunek R.o. todti jest bardziej niebieski i ma wyraźniejszy rysunek na ciele i płetwach. Podgatunek R.o. occidentalis był dłuższy czas błędnie oznaczony jako A.sjoestedti. Właściwy A.sjoestedti to „proporczykowiec niebieski”, oznaczony dawniej błędnie jako A.coeruleum.  Fot. 1 – samiec Aphyosemion lujae. 3 – samiec A.christyi. 5 – samiec A. scheeli. 5. Grupa Aphyosemion bivittatum: 5.1. A. bivittatum (Lönnsberg, 1895) – Nigeria, Kamerun (rys. 4A). Pod nazwą A.bivittatum ukrywa się właściwie cały komples bardzo zmiennych populacji, których wspólną cechą są dwa ciemne, podłużne pasy, wyraźnie widoczne u samic i młodych samców. Wiele z tych populacji osiągnęło izolację reproduktywną, mamy tu więc do czynienia z typowym nadgatunkiem (superspecies). Różne formy opisywano pod nazwami: A.bitaenianum, loennbergi, multicolor, splendopleuris itd. Rozwój ikry przebiega w sposób ciągły.  Fot. 2 – samiec A. australe. 4 – samiec A.calliurum 6 – samiec Roloffia liberiensis 6. Grupa Aphyosemion australe: 6.1. A.australe (Rachow, 1921) – Kongo aż po Gabon (rys. 4B). Grupa ta charakteryzuje się specyficznym kształtem ciała i rysunkiem płetw, a szczególnie lirowatą płetwą ogonową. Najbardziej znanym gatunkiem jest A.australe z podstawowym brązowo-czerwonym rysunkiem płetw obrzeżonych kolorem od białego po brązowo-pomarańczowy. Brzeżne promienie płetwy są wydłużone. Bardzo podobny jest A.ahli, który jednak ma płetwę odbytową i ogonową obrzeżoną kolorem jasnożółtym, natomiast w tyle ciała widoczne bywają słabe pionowe prążki. A.calliurum (synonim A.vexillifer) jest z reguły szarobrązowy do złotobrązowego z metalicznym niebieskim połyskiem, brzeżne promienie płetw są słabo wydłużone, a płetwa ogonowa jest obrzeżona matowo-żółtym kolorem. 7. Grupa Aphyosemion bualanum: 7.1. A.bualanum (Ahl, 1924) – Kamerun, Gabon, Kongo (rys. 4F). Obydwa gatunki tej grupy dają się łatwo odróżnić od innych po pionowych czerwonych prążkach na ciele i na płetwach ogonowych. A.bualanum jest metalicznie niebieski, A.exiguum jest najczęściej brązowo-pomarańczowy z zielonym połyskiem i ma bardziej owalną płetwę ogonową. 8. Grupa Aphyosemion lujae: 8.1. A.lujae (Boulenger, 1911) – środkowe Kongo (rys. 4H). Obszerna grupa gatunków z ciągłym rozwojem ikry, których wspólną cechą jest uporządkowanie czerwonych plam na ciele w 4-6 podłużnych pasów. Najbardziej znanym jest A.lujae, sprowadzony do nas z dorzecza Konga i charakteryzujący się pomarańczowym zabarwieniem płetwy ogonowej, ozdobionej w dalszej części czerwonym paskiem. W Europie rozpowszechniony jest pod nieprawidłową nazwą A.striatum, który w dorzeczu Konga nie występuje. 9. Grupa Aphyosemion labarrei: 9.1. A.labarrei Poll, 1952 – Kongo (rys. 5A). Ta poniekąd heterogeniczna grupa jest łącznikiem między grupą A.lujae a grupą A.christyi. Szczególnie A.louessense jest blisko spokrewniona z A.cameronense. Gatunki grupy A.labarrei dają się jednak odróżnić od gatunków grupy A.lujae według rysunku na ciele, który u pierwszych jest złożony z mniejszej liczby plam nieuporządkowanych w podłużne pasy. Najwyżej w tylnej części ciała widoczny jest u nich jeden nieregularny pas. A.labarrei jest metalicznie niebieski, a jego płetwa ogonowa posiada szerokie ciemnoczerwone obrzeżenie. A.louessense jest bardziej zielony i nie ma tak wyraźnie obrzeżonej płetwy ogonowej.  Fot. 7 – dojrzała para Aphyosemion gardneri (wyżej samica niżej samiec) 9 – samiec Aphyosemion filamentosum, 11 – samiec Aphyosemion walkeri, 10. Grupa Aphyosemion christyi: 10.1. A.christyi (Boulenger, 1915) – środkowe Kongo (rys. 5D). Ta charakterystyczna grupa proporczykowców z dorzecza Konga często występuje pod nazwą kompleksu „A.elegans„. A.elegans jest jednak akwarystom praktycznie nieznany. Najbardziej znanym gatunkiem jest A.christyi (dawniej A.schoutedeni), stanowiący nadgatunek złożony z licznych, bardzo zmiennych i genetycznie izolowanych, ale morfologicznie trudnych do scharakteryzowania populacji. A.christyi ma wyraźny metaliczno niebieski połysk na ciele i długie wyciągnięte płetwy, czym różni się od A.cognatum, który ma barwę podstawową ciemnoczerwoną, a brzeżne promienie płetw mało wyciągnięte. Cechą charakterystyczną większości form tej grupy (z nielicznymi wyjątkami) jest duża liczba czerwonych plamek, którymi usiane jest gęsto ciało i płetwy samców. Rozwój ikry jest ciągły.  Fot. 8 – dojrzała para Roloffia occidentalis occidentalis (wyżej samica niżej samiec) 10 – dojrzała para Aphyosemion cinnamomeum (u góry samica na dole samiec) 12 – samiec Aphyoseinion sjoestedti 11. Grupa Aphyosemion cinnamomeum: 11.1. A.cinnamomeum Clausen, 1963 – Kamerun (rys. 4M). Monotypowa grupa obejmująca jeden gatunek, lecz ze względu na kształt ciała zajmuje izolowaną pozycję. Występuje na dość ograniczonym obszarze w górach Kamerunu i nie jest podobny do żadnego gatunku z obu omawianych rodzajów. Ciało jego ma barwę cynamonowo-brunatną, a zaokrąglone płetwy są niebieskie i mają szeroki złoty brzeg. Należy do ekologicznej grupy przejściowej, wyciera się do zalanego wodą torfu na dnie zbiornika. Narybek lęgnie się z reguły po 3-4 tygodniach. 12. Grupa Aphyosemion gardneri: 12.1. A.gardneri (Boulenger, 1911) – Nigeria i zachodni Kamerun A.gardneri jest nadgatunkiem złożonym z bardzo zmiennych populacji. Dwie znane formy tego nadgatunku; niebieska z jasnożółto obrzeżonymi płetwami (nazwana błędnie A.calliurum alhi, fot. 5G) i zielono-niebieska bez żółtego obrzeżenia płetw (nazwana błędnie A.calliurum calliurum, fot. 5H) należą do najłatwiejszych w hodowli i najbardziej rozpowszechnionych proporczykowców. W roku 1963 Clausen opisał te ryby jako A.nigerianum. Według Scheela A.nigerianum jest jednak synonimem A.gardneri. Także A.scheeli jest akwarystom znany pod nazwą handlową A.burundi. Ma najczęściej szmaragdowozielone zabarwienie ciała, płetwę ogonową z węższym żółtym obrzeżeniem, któremu towarzyszy czerwony pas jedynie na dole (u góry nie występuje), co różni go od podobnej formy A.gardneri. A.ndianum wyróżnia się rysunkiem płetwy ogonowej, A.mirabilis charakteryzuje się błyszczącymi, jasnymi plamami na ciele i płetwach. Pozostałe dwa gatunki mają wiele wspólnych cech z A.louessense z dorzecza Konga. Gatunki A.gardneri należą do ekologicznej grupy pośredniej. Wycierają się zarówno w roślinnach jak i na dnie, które nie powinno wysychać. Czas rozwoju ikry jest bardzo różny (2-7 tygodni), co można obserwować zwłaszcza u A.gardneri. Ikra pozostałych gatunków rozwija się przeważnie ciągle. 13. Grupa Aphyosemion walkeri. 13.1. A.walkeri (Boulenger, 1911} – Ghana, Wybrzeże Kości Słoniowej (rys. 5M). A.walkeri jest jedynym gatunkiem rodzaju Aphyosemion, który występuje na zachód od Togo. Charakteryzuje się jesnopomarańczową barwą płetwy ogonowej, która ma środkowe promienie przedłużone, a pośrodku jej występuje czerwony rysunek i jest u góry oraz na dole czerwono obrzeżona. Gatunek ten jest także znany pod nazwą A.spurelli i odwrotnie, pod nazwą A.”ghana” długo wykazywana była Fundulosoma thierryi, która jednak nie należy do rodzaju Aphyosemion. Na swobodzie wyciera się A.walkeri na dnie, a ikra rozwija się z przerwą. Ikra stale zalana wodą może w wyższych temperaturach (23°C) rozwinąć się już po 3 tygodniach. Jednak ten sposób wychowu narybku nie wpływa dobrze na wartość potomstwa. 14. Grupa Aphyosemion filamentosum: 14.1. A.filamentosum (Meinken, 1933) – Nigeria, (rys. 6A,B). A.filamentosum i A.arnoldi tworzą grupę małych gatunków, charakteryzujących się metaliczno niebieskim kolorem oraz kształtem ciała i płetw. Typowa dla nich jest prostokątna, długa płetwa grzbietowa A.filamentosum ma w dolnej części płetwy ogonowej czerwony pas, który może jednak w niektórych populacjach nie występować. 15. Grupa Aphyosemion gulare: 15.1. A.gulare (Boulenger, 1901) – południowa Nigeria (rys. 6D). Także A.gulare jest nadgatunkiem złożonym ze zmiennych populacji. Zajmuje on pośrednie miejsce między grupami A.filamentosum i A.sjoestedti i jest szczególnie bliski grupie A.filamentosum. Jako oddzielną monotypową grupę wyodrębniono go dlatego, że różni się od drobnych gatunków grupy A.filamentosum swymi dużymi rozmiarami. (A.filamentosum dorasta do 4 cm. A.gulare 8 cm) i wykazuje już typowe cechy gatunku wycierającego się na dnie, z przerwanym rozwojem ikry. Z A.sjoestedti nie jest tak blisko spokrewniony, jak się zwykle sądziło. W akwarystyce jest znany także pod nazwą A.beauforti A.falax jest również synonimem tego gatunku. 6. Grupa Aphyoseinion sjoestedti:; 16.1. A.sjoestedti (Lonnberg, 1895) – południowa Nigeria, zachodni Kamerun (rys. 6E). Jest to największy z proporczykowców (12 cm), znany pod nazwą proporczykowca niebieskiego, charakteryzujący się swoją wielkością i trzyczęściową, pstrokątną płetwę ogonową. Długo był błędnie określany jako A.caeruleum, albo A.gulare coeruleum. Ma oczywiście także charakter nadgatunku. Wyciera się na dnie, rozwój ikry z diafazą. LITERATURA Foersch W., 1968: Neue Aphyosemionarten aus Kamerun, DATZ, 12, slr. 365-369 (Tłum. H. Skrzyński). Redakcji czechosłowackiego czasopisma akwarystycznego „akvarium a terarium” dziękujemy za zezwolenie na przedruk i przekazanie materiałów ilustracyjnych. Redakcja |

